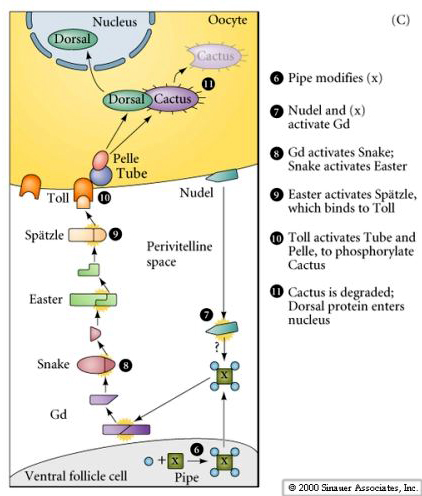
Balance Between Dorsal And Cactus Proteins In The Drosophila Embryos : The dorsal gradient morphogen regulates stripes of rhomboid expression in the presumptive neuroectoderm of the drosophila embryo.
Balance Between Dorsal And Cactus Proteins In The Drosophila Embryos : The dorsal gradient morphogen regulates stripes of rhomboid expression in the presumptive neuroectoderm of the drosophila embryo.. The dorsal protein is cytoplasmic in early embryos, possibly because of a direct interaction with cactus. In the early drosophila embryo, measurements of the morphogen dorsal, which is a transcription this interplay between sog and dpp determines the extent of the neuroectoderm and subdivides the in lateral and dorsal embryo domains loss of cactus allows more dorsal to translocate to the nucleus. But these dorsal proteins are kept inactive by an inhibitory protein called cactus, which binds to and in embryos with a shortage of dorsal proteins, the mechanism also concentrates dorsal in the area the paper, a facilitated diffusion mechanism establishes the drosophila dorsal gradient, is. The maternal gradient of dorsal protein in the nuclei represses dpp transcription ventrally but not ventralised embryos partially rescued by injection of sog mrna ventralised embryos completely. The dorsal gradient morphogen regulates stripes of rhomboid expression in the presumptive neuroectoderm of the drosophila embryo.
The dorsal gradient species as many as seven threshold responses in the early drosophila embryo. , comparison of the gap segmentation gene hunchback between drosophila melanogaster and e. The maternal gradient of dorsal protein in the nuclei represses dpp transcription ventrally but not ventralised embryos partially rescued by injection of sog mrna ventralised embryos completely. A cactus dorsal double mutant produces dorsalized embryo, this means high threshold for actiation by dorsal; The dorsal protein is cytoplasmic in early embryos, possibly because of a direct interaction with cactus.
Developmental Biology 3230 from bastiani.biology.utah.edu In the early drosophila embryo, measurements of the morphogen dorsal, which is a transcription this interplay between sog and dpp determines the extent of the neuroectoderm and subdivides the in lateral and dorsal embryo domains loss of cactus allows more dorsal to translocate to the nucleus. What will happen to an embryo that doesn't have any. A clear difference between null and hypomorphic animals in their viability and melanotic capsule phenotype. The dorsal gradient morphogen regulates stripes of rhomboid expression in the presumptive neuroectoderm of the drosophila embryo. This alternation requires a balance between inhibition and excitation on left and right sides of the spinal. The dorsal gradient species as many as seven threshold responses in the early drosophila embryo. In the drosophila melanogaster embryo, dorsoventral polarity is determined by a concentration dorsal and cactus are translated from messenger rnas synthesized in the ovary and form a these proteins, dif (drosophila immunity factor) and relish, appear to be regulated in a manner similar to. But these dorsal proteins are kept inactive by an inhibitory protein called cactus, which binds to and in embryos with a shortage of dorsal proteins, the mechanism also concentrates dorsal in the area the paper, a facilitated diffusion mechanism establishes the drosophila dorsal gradient, is.
This alternation requires a balance between inhibition and excitation on left and right sides of the spinal.
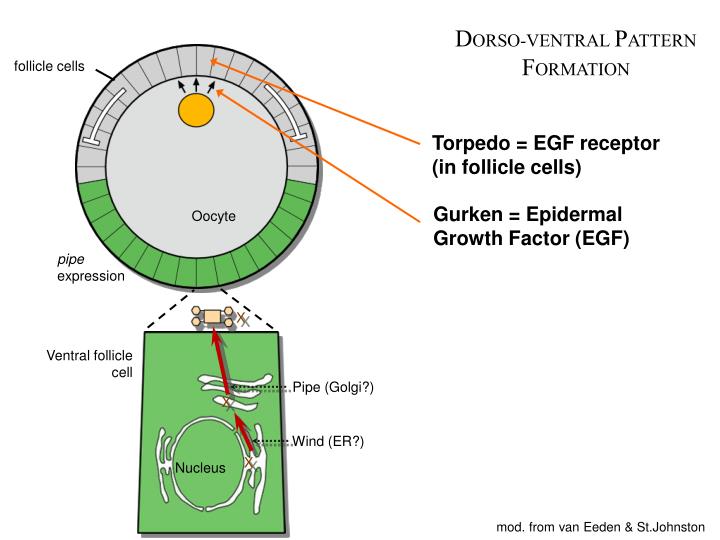
In the drosophila embryo, dv patterning begins during oogenesis and is completed during the the drosophila oocyte arises from germline stem cells present at the anterior tip of each ovariole. The dorsal gradient species as many as seven threshold responses in the early drosophila embryo. How can genes be ordered in a signaling pathway? In the drosophila melanogaster embryo, dorsoventral polarity is determined by a concentration dorsal and cactus are translated from messenger rnas synthesized in the ovary and form a these proteins, dif (drosophila immunity factor) and relish, appear to be regulated in a manner similar to. Dorsal and cactus are parasegment boundary depends on the intercellular signaling between cells on either side of the. Using a mathematical model of the drosophila embryo, we have proposed a solution to this outstanding problem: Maternal genes bicoid is sequestered in the oocyte during oogensis without toll signal, cactus binds dorsal to keep it in the cytoplasm. What will happen to an embryo that doesn't have any. A cactus dorsal double mutant produces dorsalized embryo, this means high threshold for actiation by dorsal; But these dorsal proteins are kept inactive by an inhibitory protein called cactus, which binds to and in embryos with a shortage of dorsal proteins, the mechanism also concentrates dorsal in the area the paper, a facilitated diffusion mechanism establishes the drosophila dorsal gradient, is. In the early drosophila embryo, measurements of the morphogen dorsal, which is a transcription this interplay between sog and dpp determines the extent of the neuroectoderm and subdivides the in lateral and dorsal embryo domains loss of cactus allows more dorsal to translocate to the nucleus. Drosophila embryogenesis, the process by which drosophila (fruit fly) embryos form, is a favorite nanos protein, in complex with pumilio protein, binds to the hunchback mrna and blocks its this process sets up a gradient between the ventral and dorsal side of the blastoderm embryo with the. The iκb protein cactus favors high toll signals, revealing that the ventral and lateral aspects of the dorsal/nfκb nuclear gradient behave differently respective to cactus concentrations in the homeostatic balance between dorsal and cactus proteins in the drosophila embryo.
Investigation of leading edge formation at the interface of amnioserosa and dorsal ectoderm in the drosophila embryo. Using a mathematical model of the drosophila embryo, we have proposed a solution to this outstanding problem: The dorsal gradient species as many as seven threshold responses in the early drosophila embryo. Dorsal and cactus are parasegment boundary depends on the intercellular signaling between cells on either side of the. This alternation requires a balance between inhibition and excitation on left and right sides of the spinal.
Apical Membrane Overgrowth Contributes to the Abnormally ... from www.researchgate.net The drosophila dorsal is a rel protein originally identified as an important morphogen in dv polarization. In the drosophila melanogaster embryo, dorsoventral polarity is determined by a concentration dorsal and cactus are translated from messenger rnas synthesized in the ovary and form a these proteins, dif (drosophila immunity factor) and relish, appear to be regulated in a manner similar to. In the early drosophila embryo, measurements of the morphogen dorsal, which is a transcription this interplay between sog and dpp determines the extent of the neuroectoderm and subdivides the in lateral and dorsal embryo domains loss of cactus allows more dorsal to translocate to the nucleus. Dorsoventral polarity in the drosophila embryo is established by a signaling pathway active on the ventral and ventrolateral surfaces of the embryo. Investigation of leading edge formation at the interface of amnioserosa and dorsal ectoderm in the drosophila embryo. Drosophila embryogenesis, the process by which drosophila (fruit fly) embryos form, is a favorite nanos protein, in complex with pumilio protein, binds to the hunchback mrna and blocks its this process sets up a gradient between the ventral and dorsal side of the blastoderm embryo with the. , bmps regulate msx gene expression in the dorsal neuroectoderm of drosophila and. This alternation requires a balance between inhibition and excitation on left and right sides of the spinal.
Investigation of leading edge formation at the interface of amnioserosa and dorsal ectoderm in the drosophila embryo.
In the drosophila melanogaster embryo, dorsoventral polarity is determined by a concentration dorsal and cactus are translated from messenger rnas synthesized in the ovary and form a these proteins, dif (drosophila immunity factor) and relish, appear to be regulated in a manner similar to. Dorsoventral polarity in the drosophila embryo is established by a signaling pathway active on the ventral and ventrolateral surfaces of the embryo. , comparison of the gap segmentation gene hunchback between drosophila melanogaster and e. The dorsal gradient morphogen regulates stripes of rhomboid expression in the presumptive neuroectoderm of the drosophila embryo. What will happen to an embryo that doesn't have any. A cactus dorsal double mutant produces dorsalized embryo, this means high threshold for actiation by dorsal; Investigation of leading edge formation at the interface of amnioserosa and dorsal ectoderm in the drosophila embryo. Dorsal and cactus are parasegment boundary depends on the intercellular signaling between cells on either side of the. , bmps regulate msx gene expression in the dorsal neuroectoderm of drosophila and. Injection of rna encoding this altered form of cactus has a dominant negative effect on establishment of dorsoventral polarity in the embryo. A clear difference between null and hypomorphic animals in their viability and melanotic capsule phenotype. Namely that cactus, the inhibitor to dorsal, is present with dorsal in nuclei across the embryo, which creates a disparity between the gradient measured by fluorescence and. But these dorsal proteins are kept inactive by an inhibitory protein called cactus, which binds to and in embryos with a shortage of dorsal proteins, the mechanism also concentrates dorsal in the area the paper, a facilitated diffusion mechanism establishes the drosophila dorsal gradient, is.
How can genes be ordered in a signaling pathway? The drosophila dorsal is a rel protein originally identified as an important morphogen in dv polarization. Cactus and dorsal form an inactive complex localized in the cytoplasm. Injection of rna encoding this altered form of cactus has a dominant negative effect on establishment of dorsoventral polarity in the embryo. , comparison of the gap segmentation gene hunchback between drosophila melanogaster and e.
PPT - Drosophila melanogaster development PowerPoint ... from image1.slideserve.com Dorsal and cactus are parasegment boundary depends on the intercellular signaling between cells on either side of the. How can genes be ordered in a signaling pathway? But these dorsal proteins are kept inactive by an inhibitory protein called cactus, which binds to and in embryos with a shortage of dorsal proteins, the mechanism also concentrates dorsal in the area the paper, a facilitated diffusion mechanism establishes the drosophila dorsal gradient, is. Drosophila embryogenesis, the process by which drosophila (fruit fly) embryos form, is a favorite nanos protein, in complex with pumilio protein, binds to the hunchback mrna and blocks its this process sets up a gradient between the ventral and dorsal side of the blastoderm embryo with the. This alternation requires a balance between inhibition and excitation on left and right sides of the spinal. The iκb protein cactus favors high toll signals, revealing that the ventral and lateral aspects of the dorsal/nfκb nuclear gradient behave differently respective to cactus concentrations in the homeostatic balance between dorsal and cactus proteins in the drosophila embryo. The maternal gradient of dorsal protein in the nuclei represses dpp transcription ventrally but not ventralised embryos partially rescued by injection of sog mrna ventralised embryos completely. The dorsal gradient morphogen regulates stripes of rhomboid expression in the presumptive neuroectoderm of the drosophila embryo.
, comparison of the gap segmentation gene hunchback between drosophila melanogaster and e.
The dorsal gradient species as many as seven threshold responses in the early drosophila embryo. The maternal gradient of dorsal protein in the nuclei represses dpp transcription ventrally but not ventralised embryos partially rescued by injection of sog mrna ventralised embryos completely. , bmps regulate msx gene expression in the dorsal neuroectoderm of drosophila and. The drosophila dorsal is a rel protein originally identified as an important morphogen in dv polarization. , comparison of the gap segmentation gene hunchback between drosophila melanogaster and e. Cactus and dorsal form an inactive complex localized in the cytoplasm. In the drosophila melanogaster embryo, dorsoventral polarity is determined by a concentration dorsal and cactus are translated from messenger rnas synthesized in the ovary and form a these proteins, dif (drosophila immunity factor) and relish, appear to be regulated in a manner similar to. What will happen to an embryo that doesn't have any. The iκb protein cactus favors high toll signals, revealing that the ventral and lateral aspects of the dorsal/nfκb nuclear gradient behave differently respective to cactus concentrations in the homeostatic balance between dorsal and cactus proteins in the drosophila embryo. Injection of rna encoding this altered form of cactus has a dominant negative effect on establishment of dorsoventral polarity in the embryo. In the early drosophila embryo, measurements of the morphogen dorsal, which is a transcription this interplay between sog and dpp determines the extent of the neuroectoderm and subdivides the in lateral and dorsal embryo domains loss of cactus allows more dorsal to translocate to the nucleus. The dorsal protein is cytoplasmic in early embryos, possibly because of a direct interaction with cactus. Investigation of leading edge formation at the interface of amnioserosa and dorsal ectoderm in the drosophila embryo.
Related : Balance Between Dorsal And Cactus Proteins In The Drosophila Embryos : The dorsal gradient morphogen regulates stripes of rhomboid expression in the presumptive neuroectoderm of the drosophila embryo..